Emésztés
Az oldal jelenlegi verzióját még nem ellenőrizték tapasztalt közreműködők, és jelentősen eltérhet a 2020. december 17-én felülvizsgált verziótól ; az ellenőrzéshez 31 szerkesztés szükséges .Emésztés - élelmiszer kémiai (főleg enzimatikus), esetenként mechanikai feldolgozása - olyan folyamatok összessége, amelyek biztosítják a tápanyagok felszívódásra és az anyagcserében való részvételre alkalmas komponensekre való lebontását [1] . Az emésztés során az élelmiszer szerves makromolekulái kisebb molekulákká alakulnak, különösen az élelmiszer- biopolimerek monomerekké bomlása. Ezt a folyamatot emésztő (hidrolitikus) enzimek segítségével hajtják végre . A fenti feldolgozási folyamat után a táplálék közvetlenül a sejtek citoplazmájába szívódik fel (baktériumokban és gombákban az extracelluláris emésztés és az intracelluláris emésztés során), vagy az emésztőrendszer falain keresztül a testnedvekbe (emberben a vérbe vagy a nyirokba).
Az emésztés biológiai jelentősége
- A nagy részecskék kisebbekre bontása szükséges a táplálék felszívódásához - a sejtmembránon keresztül a sejtek citoplazmájába történő szállításához, az intraintestinalis emésztéssel rendelkező állatoknál pedig a gyomor-bél traktus falán keresztül történő felszívódásához a szállítórendszerbe ( vér, nyirok és így tovább).
- A fehérjék, a DNS (és részben más élelmiszer-polimerek) monomerekre történő hasítása szükséges az ilyen típusú organizmusokra jellemző „saját” biomolekulák monomerekből történő későbbi szintéziséhez.
Az emésztés főbb típusai és megoszlásuk az élő szervezetek csoportjai között
Extracelluláris (távoli) emésztés
Erre a fajta emésztésre az a jellemző, hogy a kiválasztó sejtek által szintetizált enzimek az extracelluláris környezetbe kerülnek, ahol hidrolitikus hatásuk megvalósul. A hidrolitikus enzimek sejtekből az emésztőüregekbe történő szekréciója először koelenterátumokban és ctenoforokban jelent meg . A lándzsafélékben , rákfélékben , rovarokban , lábasfejűekben , zsákállatokban és húrfélékben (a lándzsa kivételével ) az extracelluláris emésztés dominál. Különösen jól szervezett állatokban és emberekben fejlesztették ki, ahol a tápanyagok kezdeti emésztését biztosítja. Ezekben a szervezetekben a szekréciós sejtek elég távol helyezkednek el az emésztőüregektől, ahol a hidrolázok működése megvalósul , ezért az extracelluláris emésztés távolinak minősül. A speciális üregekben előforduló távoli emésztést üregnek nevezik. [2]
Az extracelluláris emésztés minden heterotróf szervezetre jellemző, amelynek sejtfala van - baktériumok, archeák, gombák, húsevő növények stb. Ezzel az emésztési módszerrel az enzimek a külső környezetbe szekretálódnak, vagy a külső membránon rögzülnek (gramm-ban). negatív baktériumok) vagy a sejtfalon. A táplálék emésztése a sejten kívül történik, a keletkező monomerek a sejtmembrán transzporter fehérjék segítségével szívódnak fel .
Cavitáris emésztésAz üreges emésztés a többsejtű állatokra jellemző, amelyek gyomor-bél traktussal rendelkeznek , és ez utóbbi üregében fordul elő.
Extraintestinalis emésztésA bélrendszeren kívüli emésztés jellemző egyes állatokra, amelyeknek van bélrendszerük, de emésztőenzimeket juttatnak a zsákmány testébe, majd felszívják a félig emésztett táplálékot (a pókok és az úszóbogár lárvái jól ismert példái az ilyen állatoknak).
Membrán (parietális) emésztésA parietális emésztést a vékonybél mikrobolyhai közötti nyálkarétegben végzik, és közvetlenül a felszínükön (a glikokalixben ) gerinceseknél és néhány más állatnál.
Intracelluláris emésztés
Az intracelluláris emésztés az endocitózishoz szorosan kapcsolódó folyamat, és csak az eukarióták azon csoportjaira jellemző, amelyeknek nincs sejtfaluk (egyes protisták és a legtöbb állat). Filogenetikailag ez a fajta emésztés a legősibb, mivel a legegyszerűbb és legprimitívebb többsejtű szervezetekben, gyakran laposférgekben gyakori . Kétféle intracelluláris emésztés létezik. Az első a kis molekulák sejtmembránokon keresztül történő szállításával, majd a citoszol enzimek általi emésztésével kapcsolatos . Az intracelluláris emésztés speciális intracelluláris üregekben - emésztési vakuolákban is megtörténhet, amelyek folyamatosan jelen vannak vagy fagocitózis és pinocitózis során képződnek, és eltűnnek, miután a befogott táplálék feldarabolódik. Ez a második típusú intracelluláris emésztés a legtöbb esetben a lizoszómák részvételével jár, amelyek különféle hidrolitikus enzimek széles skáláját tartalmazzák, optimális pH-értéke 3,5-5,5, függetlenül attól, hogy melyik szervezethez tartoznak. A lizoszóma enzimek jelentősen eltérnek az extracelluláris emésztésben részt vevőktől. [2]
Az élelmiszer-részecskék vagy élelmiszer-oldatok a pericelluláris környezetben a plazmamembrán invaginációit okozzák, amelyek azután összefűződnek és a citoplazmába süllyednek, pino- és fagocita vakuolákat képezve. Ez utóbbihoz kapcsolódva a lizoszómák úgynevezett fagoszómákat alkotnak , ahol az enzimek érintkezése a megfelelő szubsztrátokkal történik. Ebben az esetben intracelluláris extraplazmatikus emésztés megy végbe, melynek fizikai-kémiai törvényszerűségei nem, vagy alig térnek el a hasi emésztés törvényeitől. Ezért a második típusú intracelluláris emésztés mikroüregként jellemezhető. Mivel a lizoszómák és vakuolák membránjának belső felülete különböző hidrolitikus enzimeket tartalmaz, a mikroüreges hidrolízist ki lehet egészíteni membránhidrolízissel. A keletkező hidrolízistermékek a fagoszóma membránon keresztül szívódnak fel. Az emésztési ciklus befejezése után a fagoszómamaradékok exocitózissal kilökődnek a sejten kívülre . A lizoszómák fontos szerepet játszanak a sejt saját struktúráinak lebontásában is, amelyeket akár a sejt, akár azon kívül táplálékként használ fel. [2]
Emésztés állatokban
A legtöbb állatnál az intraintesztinális emésztés kombinálódik az intracelluláris emésztéssel. A szivacsokban csak intracelluláris emésztés van jelen . A túlnyomórészt bélen belüli emésztés (néha extraintestinális emésztéssel kiegészítve) a rovarokra , fonálférgekre és gerincesekre jellemző . Egyes állatoknál a belek hiányoznak (szivacsok, placozoa, cnidosporidia, dicyemids és orthonectids, pogonoforok, galandférgek, trematoda sporociszták, a rhizocephalic rákok felnőtt stádiumai és más csoportok bizonyos fajai). Az ilyen állatok intracelluláris emésztést (szivacsok, plakozoánok), extraintestinalis külső vagy parietális emésztést (placozoánok, galandférgek) alkalmaznak, vagy endoszimbiontákkal táplálkoznak. amelyek intracellulárisan emészthetők (pogonoforok, egyes kagylók).
Egyes állatoknál (cnidarians, nemertodermatids és xenoturbellids, sok laposféreg) a belek vakon záródnak (csak a szájnyílás vezet bele). Ugyanakkor gyakran kialakul az izmos garat, a turbellariákban garatmirigyek nyílnak meg, amelyek emésztőenzimeket szabadítanak fel.
A legtöbb állatnál az emésztőrendszer áthalad. Tartalmazza az elülső, középső és hátsó bélt; az elülső és a hátsó bél az ektodermából, a középső bél az endodermából fejlődik ki. Az emésztőrendszer gyakran funkcionális részekre oszlik - a szájüreg, a garat, a nyelőcső, a gyomor és a belek. A puhatestűek és a legtöbb ízeltlábú emésztése magában foglalja az emésztőmirigyeket - a hepatohasnyálmirigyet, amelynek vezetékei a gyomorba nyílnak, és gyakran a nyálmirigyeket is.
Az ízeltlábúakban, az előbélben (különösen a gyomorban) néhány túlnyomórészt növényevő faj kitinszerű képződményekkel rendelkezik, amelyek a szilárd táplálék őrlésére szolgálnak. Az orális apparátust módosított végtagok alkotják.
A gerincesekben az emésztés a következő egymással összefüggő folyamatok kombinációja: az élelmiszer mechanikai és fizikai feldolgozása, mozgása az emésztőrendszeren keresztül, az élelmiszer-összetevők kémiai megsemmisítése ( hidrolízis ), amelyet a gyomor-bél traktus szekréciós funkciója valósít meg ; a szerves és szervetlen vegyületek, köztük a nyomelemek és a víz felszívódásának folyamata a vérbe és a nyirokba ; a szervezet eltávolítandó salakanyagainak kiválasztódása a gyomor-bél traktus lumenébe ; azok eltávolítása a szervezetből az emésztetlen ételmaradékokkal együtt.
A gerincesekre jellemző az intracelluláris emésztés hiánya vagy gyenge expressziója, valamint az intraintesztinális és parietális emésztés túlsúlya. Minden gerincesnél a máj és a hasnyálmirigy, a szárazföldi állatoknál a nyálmirigyek is részt vesznek az emésztésben.
Az emberi emésztési folyamat
Szájüreg
Az emberben az emésztés a szájban kezdődik , ahol az ételt megrágják. Ez a folyamat serkenti a külső elválasztású mirigyeket a nyál kiválasztására . A nyálban jelenlévő amiláz részt vesz a poliszacharidok lebontásában és a bolus - egy táplálékcsomó - képződésében , amely megkönnyíti a táplálék áthaladását a nyelőcsövön . A garat nyálkahártyájában lévő receptorok irritációja okozza a nyelési reflexet , amely a medulla oblongata és a pons varolii nyelési központjában koordinálódik . Az összehangolt nyelési aktus magában foglalja a lágy szájpadlást és a uvulát , amelyek megakadályozzák, hogy a táplálék bejusson az orrüregbe , valamint az epiglottis , amely megakadályozza, hogy a táplálék bejusson a légcsőbe .
Gyomor
A gyomor a rekeszizom alatt található a bal hypochondriumban és az epigasztrikus régióban. 3 skin van:
- Külső (hashártya)
- izomréteg
- külső réteg (hosszirányú)
- középső réteg (kör alakú);
- belső réteg (ferde).
- Belső (nyálkahártya) - nem keratinizált, hengeres hámréteggel bélelt.
Az élelmiszer a szívizomzaton keresztül jut be a gyomorba . Ott keveredik a gyomornedvvel , amelynek aktív összetevői a sósav és az emésztőenzimek :
- Pepszin - a fehérjéket aminosavakra , polipeptidekre , oligopeptidekre bontja [3] .
- Rennin (vagy kimozin ) – 1 évesnél fiatalabb gyermekeknél kapható, segíti a tejtermékek emésztését . Egy év elteltével a kimozin eltűnik, funkcióit a sósav látja el.
A gyomor parietális sejtjei szintén kiválasztják a Castle intrinsic faktorát , amely a B 12 vitamin felszívódásához szükséges .
Vékonybél
A pylorus sphincteren keresztül a táplálék a vékonybélbe jut . A vékonybél első szakasza a duodenum , ahol a táplálék összekeveredik az epével , amely a zsírok emulgeálását biztosítja a hasnyálmirigy és a vékonybél enzimei által, amelyek lebontják a szénhidrátokat ( maltóz , laktóz , szacharóz ), a fehérjéket ( tripszin és kimotripszin ). A vékonybélben a tápanyagok és vitaminok fő mennyisége a bélfalon keresztül szívódik fel.
Vastagbél
A vékonybélen való áthaladás után a táplálék bejut a vastagbélbe , amely a vakbélből , vastagbélből , szigmabélből és végbélből áll . Itt történik a víz és az elektrolitok felszívódása, és itt történik a széklet képződése is .
Az emésztés szabályozása
Az emberben az emésztés pszichofiziológiai folyamat. Ez azt jelenti, hogy a reakciók sorrendjét és sebességét befolyásolja a gyomor-bél traktus humorális képessége , a táplálék minősége és az autonóm idegrendszer állapota .
Az emésztést befolyásoló humorális képességeket a gyomor és a vékonybél nyálkahártyájának sejtjei által termelt hormonok határozzák meg . A fő emésztőhormonok a gasztrin , szekretin és kolecisztokinin . A gasztrointesztinális traktus keringési rendszerébe kerülnek, és hozzájárulnak az emésztőnedvek termelődéséhez és a táplálék mozgásához.
Az emészthetőség az élelmiszer minőségétől függ [4] :
- jelentős rosttartalom (beleértve az oldható anyagokat is) jelentősen csökkentheti a felszívódást;
- egyes élelmiszerekben található nyomelemek befolyásolják az anyagok felszívódását a vékonybélben [5] ;
- a különböző természetű zsírok különböző módon szívódnak fel. A telített állati zsírok sokkal könnyebben szívódnak fel és alakulnak emberi zsírokká, mint a többszörösen telítetlen növényi zsírok, amelyek gyakorlatilag nem vesznek részt az emberi zsír képződésében;
- a szénhidrátok, zsírok és fehérjék bélből történő felszívódása a napszaktól és az évszaktól függően némileg változik;
- a felszívódás a korábban a bélbe került termékek kémiai összetételétől függően is változik.
Az emésztés szabályozásáról is a vegetatív idegrendszer gondoskodik . A paraszimpatikus rész serkenti a szekréciót és a perisztaltikát , míg a szimpatikus rész gátolja .
Az emésztést befolyásoló hormonok és egyéb biológiailag aktív anyagok
Gastroenteropancreatic endokrin rendszer - az endokrin rendszer részlege , amelyet endokrin sejtek ( apudociták ) és peptiderg neuronok képviselnek, amelyek az emésztőrendszer különböző szerveiben szétszórtan termelnek peptid hormonokat . Ez a diffúz endokrin rendszer leginkább tanulmányozott része ( az APUD rendszer szinonimája ), és sejtjeinek körülbelül a felét foglalja magában. A gasztroenteropancreas endokrin rendszerét "az emberi test legnagyobb és legösszetettebb endokrin szervének" [6] nevezték .
A gasztrin a gyomor G-sejtjei által szintetizált hormon, amely főként a gyomor pylorus régiójában található. A gasztrin a gyomorbanspecifikus gasztrinreceptorokhoz kötődikA gyomor parietális sejtjeiben megnövekedett adenilát-cikláz aktivitás eredménye a gyomornedv , különösen a sósav szekréciójának fokozódása . A gasztrin növeli a gyomor fő sejtjeinek pepszin szekrécióját is, ami a gyomornedv savasságának növekedésével együtt,optimális pH - t biztosítva a pepszin működéséhez, elősegíti a táplálék optimális emésztését a gyomorban. Ugyanakkor a gasztrin fokozza a bikarbonátok és a nyálka szekrécióját a gyomornyálkahártyában, ezáltal védi a nyálkahártyát a sósav és a pepszin hatásaitól. A gasztrin gátolja a gyomor kiürülését, ami biztosítja, hogy a sósav és a pepszin elegendő ideig érintkezzen az élelmiszer-bolusszal az emésztés érdekében. Ezenkívül a gasztrin fokozza a prosztaglandin E termelődését a gyomornyálkahártyában, ami helyi értágulathoz, fokozott vérellátáshoz és a gyomornyálkahártya fiziológiás ödémájához, valamint a leukociták gyomornyálkahártyába történő migrációjához vezet.
A szekretin egy 27 aminosavból álló peptid hormon, amelyet a vékonybél nyálkahártyájának S-sejtjei termelnek , és részt vesz a hasnyálmirigy szekréciós aktivitásának szabályozásában . Az epesavak fokozzák a szekretintermelés stimulálását [6] . A vérbe felszívódva a szekretin eléri a hasnyálmirigyet, ahol fokozza a víz és az elektrolitok, főleg a bikarbonát szekrécióját . A hasnyálmirigy által kiválasztott lé mennyiségének növelésével a szekretin nem befolyásolja a mirigy enzimképződését. Ezt a funkciót egy másik, a vékonybél nyálkahártyájában termelődő anyag – kolecisztokinin – látja el . A szekretin biológiai meghatározása azon a képességén alapul, hogy (állatoknak intravénásan adva) növeli a lúg mennyiségét a hasnyálmirigy levében [7] . A szekretin a gyomor parietális sejtjei sósavtermelésének blokkolója [8] . A szekretin által okozott fő hatás az epe , a hasnyálmirigy utak és a Brunner-mirigyek hámja által kiváltott bikarbonáttermelés stimulálása , így a táplálékfelvétel hatására a bikarbonát szekréció akár 80%-át is biztosítják. Ezt a hatást a kolecisztokinin szekréciója közvetíti,és ez az epetermelés fokozódásához, az epehólyag és a bélösszehúzódások stimulálásához, valamint a bélnedv szekréciójának fokozásához vezet [6] .
A kolecisztokinin ( CCK ; korábban pancreozymin ) egy neuropeptid hormon, amelyet a nyombél nyálkahártyájában és a proximális jejunumban lévő I-sejtek termelnek [9] . A kolecisztokinin közvetítőként működik a szervezetben előforduló különféle folyamatokban, beleértve az emésztést is. A kolecisztokinin stimulálja az Oddi záróizom relaxációját ; növeli a máj epe áramlását; növeli a hasnyálmirigy szekrécióját; csökkenti a nyomást az eperendszerben: a pylorus összehúzódását okozza, ami gátolja az emésztett tápláléknak a nyombélbe való mozgását [10] [6] . A kolecisztokinin a gyomor parietális sejtjeinek sósavszekréciójának blokkolója [8] . A szomatosztatin egy kolecisztokinin inhibitor.
A glükóz - dependens inzulinotróp polipeptid(korábban elterjedt elnevezése: gastroinhibitory polipeptid , gastric inhibitory peptide _ _ _a jejunum része [ 6] . A secretin családhoz tartozik . A glükózfüggő inzulinotróp polipeptid egy inkretin , vagyis a bélben termelődik szájon át történő táplálékfelvétel hatására. A glükóz-dependens inzulinotróp polipeptid fő funkciója a hasnyálmirigy béta-sejtjeinek inzulin -szekréciójának serkentése a táplálékfelvétel hatására. Ezenkívül a GIP gátolja a zsírok felszívódását, gátolja a nátrium és a víz reabszorpcióját az emésztőrendszerben , gátolja a lipoprotein lipázt [6] .
A vazoaktív intestinális peptid (más néven vazoaktív intestinális polipeptid ; gyakori rövidítések VIP és VIP ) egy neuropeptid hormon, amely 28 aminosavból áll, amelyek számos szervben megtalálhatók, beleértve a beleket , az agyat és a gerincvelőt , valamint a hasnyálmirigyet [6] . A vasoaktív intestinalis peptid, a szekretin család többi peptidhormonjával ellentétben, kizárólag neurotranszmitter . Erősen serkenti a bélfal véráramlását, valamint a bél simaizomzatát [6] . Ez egy olyan inhibitor, amely gátolja a sósav kiválasztását a gyomornyálkahártya parietális sejtjeiben [ 11] . A VIP emellett serkentia gyomor fő sejtjeinek pepszinogén termelését [12] .
A motilin egy hormon, amelyet a gyomor-bél traktus nyálkahártyájának kromaffin sejtjei termelnek, elsősorban a nyombél és a jejunum .
A szomatosztatin a hasnyálmirigy Langerhans-szigeteinek delta sejtjeinek hormonja, valamint a hipotalamusz egyik. Kémiai szerkezete szerint peptid hormon. A szomatosztatin gátolja a szomatotropin-felszabadító hormon szekrécióját a hipotalamuszban , valamint a szomatotrop hormon és a pajzsmirigy-stimuláló hormon szekrécióját az agyalapi mirigy elülső részében . Ezenkívül gátolja a különböző hormonálisan aktív peptidek ésa gyomorban , a belekben , a májban és a hasnyálmirigyben termelődő szerotonin szekrécióját . Különösen csökkenti az inzulin , a glukagon , a gasztrin , a kolecisztokinin , a vazoaktív bélpeptid , az inzulinszerű növekedési faktor-1 szekrécióját .
Emésztőenzimek
Az emésztőenzimek olyan enzimcsoportok , amelyek az élelmiszerek összetett összetevőit kémiailag egyszerűbb anyagokká bontják le, amelyek ezután közvetlenül a szervezetbe szívódnak fel, vagy bejutnak a keringési rendszerbe . Tágabb értelemben az emésztőenzimek minden olyan enzimet is jelentenek, amely nagy (általában polimer) molekulákat monomerekre vagy kisebb részekre bont. Emésztőenzimek termelődnek és hatnak az emberek és állatok emésztőrendszerében . Ezenkívül a lizoszómák intracelluláris enzimei az ilyen enzimeknek tulajdoníthatók . Az emésztőenzimek fő működési helyei emberekben és állatokban a szájüreg , a gyomor és a vékonybél . Az emésztőenzimeket az emésztőszervek mirigyszövete termeli: nyálmirigyek , gyomormirigyek, máj , hasnyálmirigy és a vékonybél mirigyei. Ezenkívül az enzimatikus funkciók egy részét az obligát bélmikroflóra látja el.
Bél mikroflóra
Az emberi vastagbélben élő mikroorganizmusok emésztőenzimeket választanak ki, amelyek elősegítik bizonyos típusú élelmiszerek megemésztését:
- E. coli - elősegíti a laktóz emésztését ;
- Laktobacillusok – a laktózt és más szénhidrátokat tejsavvá alakítják .
Rovarevő növények emésztőenzimei
Proteázokat izoláltunk a Nepenthes macferlanei szekréciójából , és lipázaktivitást is kimutattunk. Fő enzime, a nepentezin szubsztrátspecifitásában a pepszinhez hasonlít [ 13] .
Lásd még
Jegyzetek
- ↑ M. S. Gilyarov. Biológiai enciklopédikus szótár. – 1986.
- ↑ 1 2 3 A. M. Ugolev. Az emésztés alakulása és a funkciók fejlődésének elvei. – 1985.
- ↑ Jakubke H.-D., Eshkayt H. Aminosavak, peptidek, fehérjék. — M .: Mir, 1985. — 289 p.
- ↑ Faleev A. V. A harmónia varázsa Archív másolat 2008. június 23-án a Wayback Machine -nél - 2006. 6. o.
- ↑ Például L. Fakambi tudományos munkái kimutatták, hogy a magas kalciumtartalmú érett sajtok megtartják a zsírok egy részét, és megakadályozzák azok felszívódását a belekben. Ennek megfelelően a zsírnak ez a része nem jut be a szervezetbe, hanem a széklettel távozik.
- ↑ 1 2 3 4 5 6 7 8 Maev I. V., Samsonov A. A. A nyombél betegségei. M. , MEDpress-inform, 2005, - 512 s, ISBN 5-98322-092-6 .
- ↑ Emésztés // Nagy Szovjet Enciklopédia : [30 kötetben] / ch. szerk. A. M. Prohorov . - 3. kiadás - M . : Szovjet Enciklopédia, 1969-1978.
- ↑ 1 2 Butov M.A., Kuznetsov P.S. Emésztőrendszeri betegségekben szenvedő betegek vizsgálata. 1. rész. Gyomorbetegségben szenvedő betegek vizsgálata Archivált 2011. szeptember 22. a Wayback Machine -nél . Tankönyv a belgyógyászati betegségek propedeutikájáról az Általános Orvostudományi Kar 3. évfolyamos hallgatói számára. Ryazan. 2007 (2,42 MB).
- ↑ Murray R, Grenner D, Mays P, Rodwell W. Human Biochemistry Archivált : 2015. január 23. . 2. kötet, p. 272.
- ↑ Yakovenko E. P., Grigoriev P. Ya., Agafonova N. A., Yakovenko A. V. A choleretic gyógyszerek helye a klinikai gyakorlatban Archív másolat 2009. augusztus 30-án a Wayback Machine -nél . Kezelő orvos. 2005, 6. sz.
- ↑ Acid Conditions in Children Archiválva : 2006. november 8. a Wayback Machine -nél . Az Orosz Orvostudományi Akadémia akadémikusa, V. A. Tabolin professzor szerkesztésében . M., 1999, 120 p.
- ↑ Rövid G. F. Az emésztőrendszer élettana. - Krasznodar: 2009. - 608 p. Az OOO BK "B csoport" kiadója. ISBN 5-93730-021-1 .
- ↑ Tökés A. Zoltán, Wang Chee Woon és Susan M. Chambers. A Nepenthes macferlanei L. Planta húsevő növény által kiválasztott emésztőenzimek, 1974, 119. kötet, 1. szám, 39-46.
| |
| |||
|---|---|---|---|---|
| ||||
| emberi emésztőrendszer | ||
|---|---|---|
| A gyomor felett | 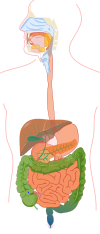 | |
| Gyomor | Pylorus | |
| Vékonybél | ||
| Kettőspont | ||
| végbélnyílás | ||
| Nagy emésztőmirigyek |
| |
| Az emésztés élettana , az emberi emésztőrendszer | |
|---|---|
| Enterális idegrendszer | |
| Enterokrin | |
| Gastroenteropancreas endokrin rendszer | |
| Enterociták | |
| biológiai folyadékok | |
| Folyamatok | |
A gyomor-bél traktus motilitása | |