Idegsejt
A neuron vagy idegsejt ( más görög νεῦρον „rost; ideg ”) egy rendkívül speciális sejt , az idegrendszer szerkezeti és funkcionális egysége . A neuron egy elektromosan gerjeszthető sejt, amelyet arra terveztek, hogy elektromos és kémiai jelek felhasználásával kívülről érkező információkat fogadjon, dolgozzon fel, tároljon, továbbítson és adjon ki.
Egy tipikus neuron egy sejttestből , dendritekből és egyetlen axonból áll . A neuronok összekapcsolódhatnak egymással, és ideghálózatokat alkothatnak . Az idegrendszer határvonalához és az információátvitel irányához viszonyítva a neuronokat receptorra (határ, kívülről veszik a jeleket, ezek alapján alakulnak ki és továbbítanak információt az idegrendszer felé), effektorokra (határ, jeleket továbbítanak az idegrendszert a külső sejtekhez) és az interkalárist (belső az idegrendszer számára).
Az idegrendszer funkcióinak összetettségét és sokszínűségét az idegsejtek, valamint az idegsejtek és az izmok és mirigyek közötti kölcsönhatás határozza meg. Ezt a kölcsönhatást az ionok segítségével továbbított különféle jelek halmaza biztosítja. Az ionok elektromos töltést ( akciós potenciált ) generálnak, amely az idegsejt testén keresztül mozog.
A neuronokat először 1837-ben Jan Purkinje fedezte fel kisagyi sejtek tanulmányozása során [1] .
A tudomány számára nagy jelentőségű volt a Golgi-módszer 1873-as feltalálása, amely lehetővé tette az egyes neuronok megfestését [2] [3] . A "neuron" ( német Neuron ) kifejezést az idegsejtek megjelölésére G. W. Waldeyer vezette be 1891-ben [4] [5] .
Az idegsejtek szerkezete
Cell törzs
Az idegsejt teste protoplazmából ( citoplazmából és sejtmagból ) áll, amelyet kívülről egy lipid kettős rétegből származó sejtmembrán ( plazmalemma , más néven neurolemma az idegsejtekben) határol . A lipidek hidrofil fejekből és hidrofób farokból állnak. A lipidek hidrofób farokba rendeződnek egymáshoz, és hidrofób réteget alkotnak. Ez a réteg csak a zsírban oldódó anyagokat (pl. oxigén és szén-dioxid) engedi át. A membránon fehérjék találhatók: a felszínen gömböcskék formájában, amelyeken poliszacharidok (glikokalix) kinövései figyelhetők meg, amelyek miatt a sejt külső irritációt észlel, valamint a membránon áthatoló integrált fehérjék, amelyekben ion található. csatornák.
A neuron egy 3-130 mikron átmérőjű testből áll. A test tartalmaz egy sejtmagot (nagyszámú nukleáris pórussal) és organellumokat (beleértve a magasan fejlett durva ER -t aktív riboszómákkal , a Golgi-készüléket ), valamint kinövéseket. Kétféle folyamat létezik: dendritek és axonok. A neuron fejlett citoszkeletonnal rendelkezik, amely behatol a folyamataiba. A citoszkeleton megtartja a sejt alakját, szálai "sínként" szolgálnak az organellumok és membránvezikulákba csomagolt anyagok (például neurotranszmitterek) szállításához. Egy neuron citoszkeletonja különböző átmérőjű fibrillákból áll: A mikrotubulusok (D = 20-30 nm) tubulin fehérjéből állnak, és a neurontól az axon mentén egészen az idegvégződésekig terjednek. A neurofilamentumok (D = 10 nm) a mikrotubulusokkal együtt biztosítják az anyagok intracelluláris transzportját. A mikrofilamentumok (D = 5 nm) aktin fehérjékből állnak, és más sejtekkel ellentétben nem tartalmaznak miozint, ami lehetetlenné teszi az összehúzódást ezekben a sejtekben, maguk a mikrofilamentumok különösen hangsúlyosak a növekvő idegi folyamatokban és a neurogliában ( Neuroglia , vagy egyszerűen csak glia ( görögül νεῦρον - rost, ideg + γλία - ragasztó) - az idegszövet segédsejtjeinek halmaza. A központi idegrendszer térfogatának kb. 40%-át teszi ki. Az agyban található gliasejtek száma kb. egyenlő a neuronok számával).
Az idegsejt testében egy fejlett szinaptikus apparátus tárul fel, az idegsejt szemcsés endoplazmatikus retikuluma bazofil módon festődik, és "tigroid" néven ismert. A tigroid behatol a dendritek kezdeti szakaszaiba, de észrevehető távolságra helyezkedik el az axon kezdetétől, ami az axon szövettani jeleként szolgál. A neuronok alakja, folyamatuk száma és funkciója különbözik. Funkciótól függően szenzitív, effektor (motoros, szekréciós) és interkalárist különböztetünk meg. A szenzoros neuronok érzékelik az ingereket, idegimpulzusokká alakítják és továbbítják az agyba. Effektor ( lat. Effectus - cselekvés) - dolgozzon ki és küldjön parancsokat a dolgozó szerveknek. Interkaláris - kapcsolatot létesít szenzoros és motoros neuronok között, részt vesz az információfeldolgozásban és a parancsok generálásában.
Az anterográd (testből) és retrográd (testbe) axontranszportot különböztetünk meg, amelyet a kinezin - dynein mechanizmus segítségével hajtanak végre (a kinezin az anterográd áramért, a dynein a retrográd áramért).
Dendritek és axon
Az axon egy neuron hosszú folyamata. Arra lett igazítva, hogy egy idegsejt ( neuroszóma ) testéből egy másik neuronba (néha ugyanarra, lásd: neuroncsapdák) vagy egy neuronból egy végrehajtó szervbe irányítson gerjesztést és információt. A dendritek egy neuron rövid és erősen elágazó folyamatai, amelyek a neuronra ható serkentő és gátló szinapszisok kialakulásának fő helyéül szolgálnak (a különböző neuronokban eltérő az axon és a dendritek hosszának aránya), és amelyek gerjesztést továbbítanak. a neuron testéhez. Egy neuronnak több dendritje és általában csak egy axonja lehet. Egy neuronnak sok (akár 20 ezer) másik neuronnal lehet kapcsolata.
A dendritek dichotóm módon osztódnak , míg az axonok kollaterálisokat eredményeznek . Az elágazó csomópontok általában mitokondriumokat tartalmaznak.
A dendriteknek nincs mielinhüvelyük , de az axonoknak igen. Az akciós potenciál (AP, „tüske”) keletkezési helye a legtöbb neuronban az axondomb ( az idegsejt trigger zónája) - az a képződés, ahol az axon a neuroszómából származik. A nyugalmi membránpotenciál ezen a helyen valamivel kisebb, vagyis a depolarizációs küszöb is kisebb. Számos kalcium- és nátriumcsatorna is részt vesz az impulzusok generálásában.
Szinapszis
A szinapszis ( görögül σύναψις , συνάπτειν - ölelés, megfogás, kezet fogni) egy speciális képződmény, amely kapcsolatot biztosít két neuron között, vagy egy idegsejt és egy jelet fogadó effektor sejt között (ingerelhető szövetek sejtjei). Két sejt közötti idegimpulzus továbbítására szolgál, szinaptikus átvitel során pedig szabályozható a jel amplitúdója és frekvenciája. Egyes szinapszisok neurondepolarizációt okoznak és serkentő hatásúak, míg mások hiperpolarizációt okoznak és gátló hatásúak. Általában egy neuron gerjesztéséhez több serkentő szinapszis stimulálása szükséges.
A kifejezést Charles Sherrington angol fiziológus vezette be 1897-ben [6] .
Az akciós potenciál keletkezésének és vezetésének mechanizmusa
1937-ben az ifjabb John Zachary megállapította, hogy a tintahal óriás axonja felhasználható az axonok elektromos tulajdonságainak tanulmányozására. A tintahal axonjait azért választották, mert sokkal nagyobbak, mint az emberiek. Ha behelyez egy elektródát az axonba, megmérheti a membránpotenciálját .
Az axon membrán feszültségvezérelt ioncsatornákat tartalmaz . Lehetővé teszik az axon számára, hogy elektromos jeleket, úgynevezett akciós potenciálokat generáljon és vezesse a testén keresztül. Ezeket a jeleket a nátrium (Na + ), kálium (K + ), klór (Cl - ), kalcium (Ca 2+ ) elektromosan töltött ionjai generálják és terjesztik.
Nyomás, nyújtás, kémiai tényezők vagy a membránpotenciál változása aktiválhatja a neuront. Ez az ioncsatornák megnyílásának köszönhető, amelyek lehetővé teszik az ionok átjutását a sejtmembránon, és ennek megfelelően megváltoztatják a membránpotenciált.
A vékony axonok kevesebb energiát és metabolikus anyagot használnak fel az akciós potenciál vezetésére, de a vastag axonok lehetővé teszik a gyorsabb levezetést.
Az akciós potenciálok gyorsabb és kevésbé energiaigényes levezetése érdekében a neuronok speciális gliasejteket használhatnak az axonok bevonására, amelyeket a központi idegrendszerben oligodendrocitáknak , vagy a perifériás idegrendszerben Schwann-sejteknek neveznek. Ezek a sejtek nem fedik le teljesen az axonokat, így az axonokon rések maradnak nyitva az extracelluláris anyagok számára. Ezekben a résekben az ioncsatornák sűrűsége megnő. Ezeket Ranvier elfogóinak hívják . Rajtuk keresztül az akciós potenciál áthalad a rések közötti elektromos mezőn.
Osztályozás
Szerkezeti osztályozás
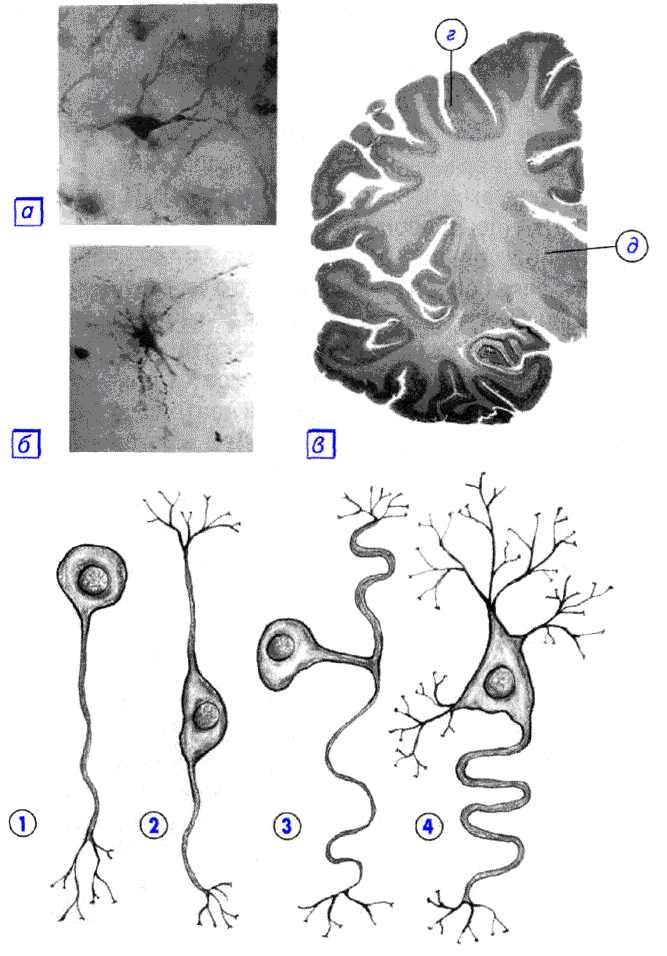
A dendritek és axonok száma és elhelyezkedése alapján a neuronokat nem-axonális, unipoláris neuronokra, pszeudounipoláris neuronokra, bipoláris neuronokra és multipoláris (sok dendrittörzs, általában efferens) neuronokra osztják [7] .
Az axonmentes neuronok a gerincvelő közelében, az intervertebralis ganglionokban csoportosuló kis sejtek , amelyek nem rendelkeznek a folyamatok dendritekre és axonokra való szétválására utaló anatómiai jelekkel. Egy sejtben minden folyamat nagyon hasonló. Az axon nélküli neuronok funkcionális célja nem teljesen ismert.
Az unipoláris neuronok - egyetlen folyamattal rendelkező neuronok, például a középagy trigeminus idegének szenzorosmagjában vannak jelen . Sok morfológus úgy véli, hogy az unipoláris neuronok nem találhatók az emberi testben és a magasabb gerincesekben.
Bipoláris neuronok - egy axonnal és egy dendrittel rendelkező neuronok, amelyek speciális érzékszervekben találhatók - a retinában, a szaglóhámban és az izzóban, a halló- és vesztibuláris ganglionokban .
A multipoláris neuronok egy axonnal és több dendrittel rendelkező neuronok. Ez a fajta idegsejtek dominálnak a központi idegrendszerben .
A pszeudo- unipoláris neuronok egyedülállóak a maguk nemében. Egy folyamat távozik a testből, amely azonnal T-alakban osztódik. Ez az egész egyetlen traktus myelinhüvellyel van borítva, és szerkezetileg egy axont képvisel, bár az egyik ág mentén a gerjesztés nem a neuron testétől, hanem a test felé halad. Szerkezetileg a dendritek ennek a (perifériális) folyamatnak a végének elágazódásai. A trigger zóna ennek az elágazásnak a kezdete (vagyis a sejttesten kívül található). Az ilyen neuronok a gerinc ganglionokban találhatók.
Funkcionális osztályozás
A reflexívben elfoglalt helyzet alapján megkülönböztetik az afferens neuronokat (érzékeny neuronokat), az efferens neuronokat (egyeseket motoros neuronoknak neveznek, néha ez nem túl pontos elnevezés az efferensek teljes csoportjára vonatkozik) és az interneuronokat ( interkaláris neuronok ).
Afferens neuronok (szenzoros, szenzoros, receptor vagy centripetális). Az ilyen típusú neuronok közé tartoznak az érzékszervek elsődleges sejtjei és a pszeudo-unipoláris sejtek, amelyekben a dendritek szabad végződésekkel rendelkeznek.
Efferens neuronok (effektor, motor, motor vagy centrifugális). Az ilyen típusú neuronok közé tartoznak a végső neuronok - ultimátum és utolsó előtti - nem ultimátum.
Asszociatív neuronok (interkaláris vagy interneuronok) - az idegsejtek egy csoportja kommunikál az efferens és az afferens között.
A szekréciós neuronok olyan neuronok, amelyek nagyon aktív anyagokat (neurohormonokat) választanak ki. Jól fejlett Golgi-komplexukkal rendelkeznek , az axon axovasális szinapszisokban végződik.
Morfológiai osztályozás
A neuronok morfológiai szerkezete változatos. A neuronok osztályozása során több elvet alkalmaznak:
- vegye figyelembe a neuron testének méretét és alakját;
- az elágazási folyamatok száma és jellege;
- az axonhossz és a speciális hüvelyek jelenléte.
A sejt alakja szerint a neuronok lehetnek gömb alakúak, szemcsések, csillag alakúak, piramis alakúak, körte alakúak, fusiformak , szabálytalanok stb. A neurontest mérete a kis szemcsés sejtekben 5 mikrontól az óriásban lévő 120-150 mikronig terjed. piramis neuronok.
A folyamatok száma szerint a következő morfológiai típusú neuronokat különböztetjük meg [8] :
- unipoláris (egy folyamattal rendelkező) neurociták, amelyek például a középagy trigeminus idegének szenzoros magjában vannak jelen;
- pszeudo-unipoláris sejtek, amelyek a gerincvelő közelében csoportosulnak az intervertebralis ganglionokban;
- bipoláris neuronok (egy axonnal és egy dendrittel rendelkeznek), amelyek speciális érzékszervekben találhatók - a retinában, a szaglóhámban és az izzóban, a halló- és vesztibuláris ganglionokban;
- multipoláris neuronok (egy axonnal és több dendrittel rendelkeznek), túlnyomórészt a központi idegrendszerben.
A neuronokat hatásuk (gátló és serkentő) és a szekretált mediátor ( acetilkolin , GABA stb.) alapján is osztályozzák, ez utóbbiakból 1985 -ben több mint 50 ismert [9] (
!)
A neuron fejlődése és növekedése
Az idegsejtek osztódásának kérdése jelenleg vita tárgya. Az egyik változat szerint az idegsejt egy kis prekurzor sejtből fejlődik ki, amely már azelőtt abbahagyja az osztódást, hogy felszabadítaná folyamatait. Először az axon kezd növekedni, majd később alakulnak ki a dendritek. Az idegsejt fejlődési folyamatának végén megvastagodás jelenik meg, amely utat nyit a környező szöveteken. Ezt a megvastagodást az idegsejt növekedési kúpjának nevezik . Az idegsejt folyamatának lapított részéből áll, sok vékony tüskével. A mikrospinulák 0,1-0,2 µm vastagok és akár 50 µm hosszúak is lehetnek; a növekedési kúp széles és lapos területe körülbelül 5 µm széles és hosszú, bár alakja változhat. A növekedési kúp mikrotüskéi közötti tereket hajtogatott membrán borítja. A mikrotüskék állandó mozgásban vannak – egyesek behúzódnak a növekedési kúpba, mások megnyúlnak, különböző irányokba térnek el, hozzáérnek az aljzathoz, és hozzátapadhatnak.
A növekedési kúp apró, néha egymással összefüggő, szabálytalan alakú hártyás hólyagokkal van tele. A membrán összehajtott területei alatt és a tüskékben összegabalyodott aktinszálak sűrű tömege található . A növekedési kúp mitokondriumokat , mikrotubulusokat és neurofilamentumokat is tartalmaz, amelyek hasonlóak az idegsejtek testében találhatóakhoz.
A mikrotubulusok és a neurofilamentumok meghosszabbítása elsősorban az újonnan szintetizált alegységek hozzáadásával történik a neuronfolyamat alapjában. Naponta körülbelül egy milliméteres sebességgel mozognak, ami megfelel az érett idegsejtek lassú axontranszportjának sebességének. Mivel az átlagos növekedési kúp előrehaladási sebessége megközelítőleg azonos, lehetséges, hogy az idegsejtek növekedése során a mikrotubulusok és neurofilamentumok sem összeépülnek, sem pedig nem pusztulnak el a távoli végén. A végén új membránanyag kerül hozzáadásra. A növekedési kúp a gyors exocitózis és endocitózis területe , amint azt az itt található számos hólyag bizonyítja. A kis membránvezikulák a neuron folyamata mentén a sejttestből a növekedési kúpba szállítódnak gyors axontranszport áramlásával. A membránanyag az idegsejt testében szintetizálódik, vezikulák formájában átkerül a növekedési kúpba, és itt exocitózissal beépül a plazmamembránba, így meghosszabbítja az idegsejt kinövését.
Az axonok és dendritek növekedését általában az idegsejtek migrációjának fázisa előzi meg, amikor az éretlen idegsejtek megtelepednek és állandó helyet találnak maguknak.
Ugyanakkor az endomitózis gyakori a neuronokban , ami szomatikus poliploidiához vezet [10] [11] .
A neuronok tulajdonságai és funkciói
Tulajdonságok- Transzmembrán potenciálkülönbség jelenléte (90 mV-ig), a külső felület elektropozitív a belső felülethez képest.
- Nagyon nagy érzékenység bizonyos vegyszerekre és elektromos áramra (általában érzékenység sokféle kémiai vegyületre, a biokémiai homeosztázis zavaraira stb.).
- A fent említett elektromos ingerlékenység , ennek hátoldala a gátlás .
- Az ingerek gerjesztési küszöbe .
- A neuroszekréció képessége , azaz speciális anyagok ( neurotranszmitterek ) szintézise és felszabadulása a környezetbe vagy a szinaptikus hasadékba.
- Magas energiafelhasználás , magas szintű energiafolyamatok, ami az oxidációhoz szükséges fő energiaforrások - glükóz és oxigén - folyamatos ellátását teszi szükségessé.
- Jelerősítés.
- Átmeneti válasz .
- Átkódolás (ritmusátalakítás).
- Tehetetlenség.
- Tanulhatóság (többnyire képernyőközponti neuronok ).
- Alkalmazkodóképesség (beleértve a ritmust - a ritmus asszimilációja).
- Moduláció (retikuláris képződés, elektromos tónus , hormonok ).
- Egy bizonyos ritmusú és szintű spontán aktivitás (beleértve a nullához nagyon közeli értéket is).
- Neuroantigenitás [12] .
- Egy adott neuron 1 összes axonterminálisának izolálása és ugyanaz: mediátor, vagy mediátorok „koktélja” (frissített Dale-elv [13] ).
- fogadó funkció. A szinapszisok érintkezési pontok; információt kapunk a receptoroktól és a neuronoktól idegimpulzus-szekvenciák (" tüske ") formájában.
- integratív funkció. Az információfeldolgozás eredményeként a neuron kimenetén jel keletkezik, amely az összes bemeneti jelből generált információt hordozza.
- karmester funkció. Az axon mentén lévő neuronból akciós potenciálok formájában jut el információ a szinapszisokhoz.
- átviteli funkció. Az idegimpulzus, miután elérte az axon végét , amely már a szinapszis szerkezetének része, egy közvetítő felszabadulását idézi elő - a gerjesztés közvetlen közvetítője egy másik neuronhoz vagy végrehajtó szervhez.
A híres amerikai neurofiziológus , Michael Graziano szerint egy adott agy idegsejtjei közötti kapcsolatok természete határozza meg ennek az agynak a lényegét és a többitől való különbségét [14] .
Lásd még
- mesterséges neuron
- Biológiai neuron modell
- neurális hálózat
- Neurofiziológia
- Tükörneuronok
- Motoneuronok
- Az állatok listája neuronok száma szerint
Jegyzetek
- ↑ Neuroszemélyiségek: Jan Purkinje, a kisagyi sejtek felfedezője . Letöltve: 2021. március 6. Az eredetiből archiválva : 2022. január 3.
- ↑ Camillo Golgi. Sulla struttura della sostanza grigia del cervelo (neopr.) // Gazzetta Medica Italiana. Lombardia. - 1873. - T. 33 . - S. 244-246 .
- ↑ Isaac Asimov . A biológia rövid története . - Ripol Classic, 2013. - 114. o.
- ↑ Jean-Pierre Changeux, Laurence Garey. Neuronális ember – Az elme biológiája . - Princeton University Press, 1997. - 28. o.
- ↑ Ugyanis a "neuron" kifejezés a következő forrásból származó "Das Nervensystem besteht aus zahlreichen, untereinander anatomisch wie genetisch nicht zusammenhängenden Nerveneinheiten (Neuronen)" idézetben fordul elő: Heinrich Wilhelm Gottfried von Waldeyer-Hartz. Ueber einige neuere Forschungen im Gebiete der Anatomie des Centralnervensystems (német) // Deutsche medicinische Wochenschrift: magazin. - 1891. - Bd. 17 , sz. 50 . — S. 1352 . - doi : 10.1055/s-0029-1206907 .
- ↑ S. A. Osipovsky; H. N. Bogolepov. Szinapszis // Big Medical Encyclopedia / szerk. B. V. Petrovszkij. - 3. kiadás - V. 23. 2017. augusztus 24-i archivált példány a Wayback Machine -nél
- ↑ I. V. Gaivaronsky. A központi idegrendszer anatómiája. Rövid tanfolyam / G. I. Nichiporuk. - 4. kiadás, átdolgozva. és további - Szentpétervár. : ELBI-SPb, 2010. - S. 3-4. — 108 p.
- ↑ lásd az ábrát. Archiválva : 2014. július 20. a Wayback Machine -nél
- ↑ Arnold J. Mandell, „From Molecular Biological Simplification to More Realistic Central Nervous System Dynamics: An Opinion”, in Psychiatry: Psychobiological Foundations of Clinical Psychiatry 3:2, JO Cavenar et al., eds. (New York: Lippincott, 1985). idézi: Gleick, James. Káosz: Új tudomány létrehozása. Penguin books, New York NY, 1987. 299. oldal. URL: [1] Archiválva : 2020. november 8. a Wayback Machine -nél
- ↑ Kirsanova, Irina Alekszandrovna. "Szomatikus poliploidia a pulmonalis haslábúak központi idegrendszerében." (2003).
- ↑ Yu. A. Modern Aspects of Somatic Polyploidy (On the First All-Union Symposium on Somatic Polyploidy in Animals, Jereván, 1971. november 10-12.) - 1972. - T. 25. - Nem. 1. - S. 112-115 .
- ↑ Ashmarin, Igor Petrovics. Az emlékezet biokémiájának rejtélyei és kinyilatkoztatásai. - Leningrád Kiadó. egyetem, 1975.
- ↑ Osborne, N. N. Dale alapelve és kommunikációja a neuronok között / szerk. NN Osborne // Anglia: Pergamon Press, 1983. - 204 p. - Hozzáférési mód: DOI: 10.1016/C2013-0-03680-8
- ↑ Graziano, 2021 , 8. fejezet.
{kind=link}
{kind=link}
Irodalom
- Polyakov G. I. Az agy idegrendszerének elveiről, M: Moszkvai Állami Egyetem, 1965
- Kositsyn N. S. Dendritek és axodendrites kapcsolatok mikrostruktúrája a központi idegrendszerben. M.: Nauka, 1976, 197 p.
- Nemechek S. et al., Introduction to Neurobiology, Avicennum: Prága, 1978, 400 pp.
- Bloom F., Leizerson A., Hofstadter L. Agy, elme és viselkedés
- Brain (cikkgyűjtemény: D. Hubel, C. Stevens, E. Kandel és mások – a Scientific American száma (1979. szeptember)). M.: Mir, 1980
- Savelyeva-Novosyolova N. A., Savelyev A. V. Egy eszköz egy neuron modellezésére. A. s. 1436720, 1988. sz
- Savelyev A. V. Az idegrendszer dinamikus tulajdonságainak variációinak forrásai szinaptikus szinten // Journal of Artificial Intelligence, National Academy of Sciences of Ukraine. - Donyeck, Ukrajna, 2006. - 4. sz . - S. 323-338 .
- Michael Graziano . A tudat tudománya. A szubjektív tapasztalat modern elmélete = Michael SA Graziano. A tudat újragondolása: a szubjektív tapasztalat tudományos elmélete. - M. : Alpina non-fiction, 2021. - 254 p. — (A Politechnika könyvei). - ISBN 978-5-00139-208-8 .
Linkek
- RF-PSTH neuromodel (a receptív mező (RP) szerkezetét és a PSTH kimeneti neurális jelet szimulálja)
- Népszerű tudományos film "Idegsejt" a YouTube -on
- Mi az a neuron? | Tudományos cikk fordítása C. George Boeree
| | ||||
|---|---|---|---|---|
| Szótárak és enciklopédiák |
| |||
| ||||